Written by Cody Cobb
 As a molecular biologist, most of my work is done on a bench at or below room temperature. I can count on one hand the number of times I’ve been to a research field because I have more than two fingers. I’ve never taken a course in ecology, and I’ve rarely dealt with full, intact organisms. It is with just such a background that I absorbed a talk by Allison Snow at Rutgers ten days ago.
As a molecular biologist, most of my work is done on a bench at or below room temperature. I can count on one hand the number of times I’ve been to a research field because I have more than two fingers. I’ve never taken a course in ecology, and I’ve rarely dealt with full, intact organisms. It is with just such a background that I absorbed a talk by Allison Snow at Rutgers ten days ago.
Snow* is an evolutionary biologist and ecologist who’s been running an interesting experiment on wild radishes for more than a decade now. In the 90’s, when transgenic crops like Bt corn and Roundup Ready soybean were beginning to dominant the market (and the landscape), there were concerns that wild relatives would incorporate the transgenes and spread as superweeds. Corn and soybean, with their lack of compatible relatives in the US, are exempt from this concern. However, as more and more transgenic crops with compatible relatives come down the pipeline (and with some, like canola, already here) there needs to be some hard data on just how easily transgenes can persist and spread in wild populations.

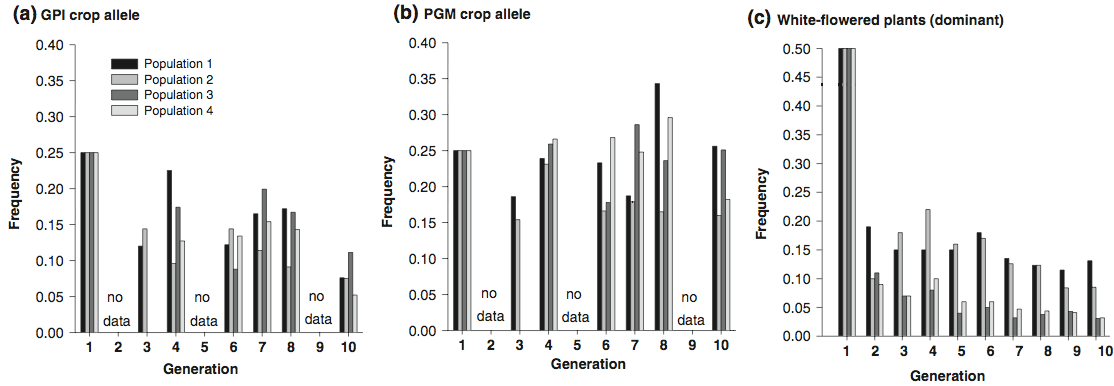

So what kind of effects in the wild can we expect to see from transgenes based on this study? From the paper’s conclusion we’re warned:
Clearly, crop alleles can persist for many generations following a single hybridization event, and crop-wild hybrids may recover wild-type fitness in later generations. Thus, beneficial or neutral transgenes that recombine independently of deleterious crop alleles may spread and persist indefinitely (Snow et al., 2010).
A relevant example is her 2003 study on Bt sunflower, which found that the Bt transgene in cultivated sunflower (Helianthus annuus), when crossed into wild sunflower (also Helianthus annuus), allows each plant to produce, on average, 55% more seeds relative to non-transgenic controls under field conditions (Snow et al., 2003). Rather frustratingly, follow-up work was halted when the companies sponsoring the study — Pioneer Hi-Bred International and Dow AgroSciences — refused to allow further access to the transgene or the seeds since they decided not to sell Bt sunflowers anyway.

With all this in mind, what are some steps we genetic manipulators and tamperers can take to lower the risk of transgene flow into wild relatives? One thought is to link the transgene of interest with another gene that’s deleterious in the wild but tolerated or even desirable in agricultural situations. Better yet, find two such genes and flank the transgene. Creating such a construct would require a lot more work, not to mention the difficulty of finding appropriate crop-tolerant-but-wild-harmful genes. But then, it’s just a thought. What are yours?
*Astute readers of Tomorrow’s Table might recognize the name from a parenthetical citation on page 110: “For this reason, some ecologists see the application of GE as a way to spare even more land from destruction by enhancing yields (Qaim and Zilberman 2003; Snow et al. 2005).”
![]() Snow, A., Pilson, D., Rieseberg, L., Paulsen, M., Pleskac, N., Reagon, M., Wolf, D., & Selbo, S. (2003). A Bt transgene reduces herbivory and enhances fecundity in wild sunflowers. Ecological Applications, 13 (2), 279-286 DOI: 10.1890/1051-0761(2003)013[0279:ABTRHA]2.0.CO;2
Snow, A., Pilson, D., Rieseberg, L., Paulsen, M., Pleskac, N., Reagon, M., Wolf, D., & Selbo, S. (2003). A Bt transgene reduces herbivory and enhances fecundity in wild sunflowers. Ecological Applications, 13 (2), 279-286 DOI: 10.1890/1051-0761(2003)013[0279:ABTRHA]2.0.CO;2
Snow AA, Culley TM, Campbell LG, Sweeney PM, Hegde SG, & Ellstrand NC (2010). Long-term persistence of crop alleles in weedy populations of wild radish (Raphanus raphanistrum). The New phytologist PMID: 20122132
Written by Guest Expert
Cody Cobb is a brand new doctor, recovering plant biologist, and photography enthusiast. He received his MD at Texas A&M Health Science Center College of Medicine. Prior to medical school, he studied plant biology and plant pathology at Rutgers University.


Great post. I’ve seen Allison Snow’s research come up many times, including her involvement in a study that didn’t find transgenes in Mexican maize. I wonder what effects those cultivation-specific genes have on the genetic diversity of the population. One of the chief objections is that introducing a transgene will crowd out genetic diversity in wild and landrace populations. I don’t give that argument much credence, however I wonder if Snow has looked into that aspect of the introgression going on in her population?
Oh yeah, the paycheck, just send me your account number and I’ll plug you into that “filthy Monsanto lucre.”
Actually, she published a short perspective last year [PDF] on more recent work that showed, in fact, there are transgenes in Mexican maize. Apparently you can’t cultivate GE corn in Mexico, but you can import it — fertile seeds and all. Seems rather silly to me, but then so many things do…