Whether glyphosate exposure causes health problems through transgenerational epigenetic inheritance is an important research question. The original EPA reference dose is based on a transgenerational phenotype, even though this result has been determined to “be spurious and unrelated to treatment since more extensive evaluations in subsequent reproduction studies conducted at much higher doses did not replicate the offspring effects” (as explained in the draft human health assessment for glyphosate).
In this series, we address questions about transgenerational inheritance and epigenetics in general, and this glyphosate study in particular.
Transgenerational inheritance is the concept that traits can be passed on from parent to great-grandchildren. In the context of toxicology, this hypothesis can be described as “ancestral environmental exposures to non-mutagenic agents can exert effects in unexposed descendants.” If you imagine a person being exposed to some substance, their reproductive cells are exposed so their children are also exposed (intergenerational inheritance). If that person is a pregnant female, the reproductive cells of their offspring are exposed so the grandchildren are also exposed (multigenerational inheritance). Thus, true transgenerational inheritance can only be observed in the great-grandchildren’s generation (transgenerational inheritance).
This graphic was originally published in a post on transgenerational exposure in the context of trauma and the Holocaust here.
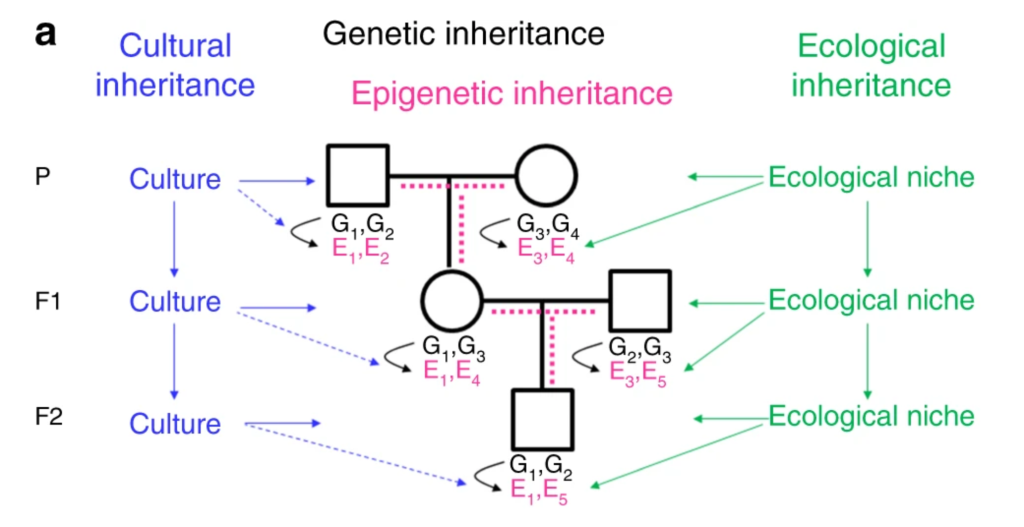
Transgenerational inheritance systems. a Offspring inherit from their parents genes (black), the environment (green) and culture (blue). Genes and the environment affect the epigenome (magenta) and the phenotype22. Culture also affects the phenotype, but at present there is no evidence for a direct effect of culture on the epigenome (broken blue lines). It is a matter of debate, how much epigenetic information is inherited through the germline (broken magenta lines). G genetic variant, E epigenetic variant.
Epigenetic inheritance
The focus of the paper under discussion is the epigenetic mechanisms through the germline, or transgenerational epigenetic inheritance. In any experiment of transgenerational inheritance, it is critical to use a careful study design to separate the epigenetic piece from the other mechanisms.
Epigenetics can be defined as: “the processes and marks on or around the DNA processes that control the activity of the genome and can be mitotically and/or meiotically inherited.” It encompasses a set of mechanisms that regulate gene expression and that can be inherited from cell to cell within an organism. Sometimes, if they occur in germline cells, these mechanisms may also be inherited from parent to offspring. Epigenetic mechanisms can also be sensitive to environmental inputs. Because they can be modified by the environment and may be inherited from parent to offspring, epigenetic mechanisms are a prime candidate for mediating transgenerational inheritance.
Epigenetics generally refers to four mechanisms.
Cytosine modifications: These are direct covalent modifications of cytosines in the DNA sequences, including DNA methylation, which is measured in the paper under discussion.
Histone modifications: Histones are proteins that, with DNA, form chromatin and make up chromosomes. Each histone has a tail that can be covalently modified.
Non-coding RNAs: These functional RNAs that are not translated into protein and are involved in many cellular processes, including regulation of the epigenome.
Long range chromatin interactions: This refers to the 3D arrangement of DNA and chromosomes within cells. In addition to the packing of DNA into chromosomes by histones, chromosomes interact with themselves and with other chromosomes to form functional domains.
These four mechanisms do not exist in isolation. They form a network of interacting mechanisms that all work together to affect gene expression. For an overview on these mechanisms of epigenetics, please visit the “Intro to Epigenetics” series at Mommy, PhD.
Transgenerational epigenetic inheritance is well documented in plants and the commonly used model organisms, such as C. elegans (roundworms) and D. melanogaster (fruit flies). However, whether transgenerational inheritance occurs in mammals is still unclear.
Does transgenerational epigenetic inheritance occur in humans?
The existence of transgenerational epigenetic inheritance remains unclear in mammals. There are a few reasons why this is hard to answer.
First, humans are complicated. When we have evidence of transgenerational inheritance of a trait in people, it is nearly impossible to separate the cultural and ecological effects from the epigenetic effects to definitively say if that inheritance occurs partly or exclusively through a biological mechanism. In humans, exposures are rarely isolated to the original generation only, making it extremely difficult to separate out true transgenerational effections. In addition, even when exposures are isolated, those exposures often produce differences that have their own effects. In the example of the Holocaust, it is difficult to separate the effects of trauma from living through the Holocaust on offspring from the effect of having a parent who lived through the Holocaust on offspring.
In order to determine if transgenerational inheritance occurs, scientists must stop the exposure in the original generation to isolate the exposure. While this can be done in model organisms in the lab, exposures are rarely isolated to a single generation in humans. Even when they are, the genetic, ecological and cultural confounders are so complex that it is extraordinarily difficult to conclusively identify transgenerational epigenetic inheritance in humans.
Second, experimental design is extremely complicated. We can use model organisms (such as mice or rats) to control for some of these factors to determine if transgenerational epigenetic inheritance occurs in mammals. However, when properly designed, these experiments are extremely complicated, expensive, and time-consuming, as described in A guide to designing germline-dependent epigenetic inheritance experiments in mammals. These experiments can be done, but at this point in time, very few studies are properly designed to actually be able to answer this question. We will discuss this in more detail below when we get to the methods of the paper under discussion.
Third, germline reprogramming clears (erases) many epigenetic marks twice during mammalian development. First, DNA methylation marks are cleared during germ-cell development. There is a second wave of demethylation following fertilization; the timing of this demethylation and the reestablishment of methylation patterns is different for maternal and paternal chromosomes. A subset of genes (mostly imprinted genes) do not undergo this second wave of demethylation and are more sensitive to environmental regulation. Thus, only a subset of the genome could be undergoing translational epigenetic inheritance. While research in the area is still evolving, it is clear that more of the genome than previously thought is protected from this second wave of demethylation. But transgenerational epigenetic inheritance seems unlikely to be a genome-wide phenomenon.
State of the transgenerational inheritance science
This 2018 state of the science report on transgenerational inheritance from the National Toxicology Program cites 21 papers from the lab that published the current glyphosate study. It summarizes the weaknesses of the existing evidence and underscores the need for well-designed studies.
“In conclusion, a broad range of exposures and outcomes have been reported to support transgenerational inheritance of health effects. Over 80 different agents have been tested in a transgenerational experimental design; and this state of the science review collected and categorized the literature into a systematic evidence map for transgenerational inheritance by broad health effect categories, exposures, and evidence streams. This scoping review and evidence map identifies serious limitations in the available bodies of evidence to support a systematic review for reaching hazard conclusions or even rating certainty in the bodies of evidence under evidence to decision frameworks such as the Grading of Recommendations Assessment, Development and Evaluation (GRADE) approach.”
This report includes assessments of potential bias in the studies that do exist. The images below show a summary of their assessment of bias in animals studies (top) and specifically in animal studies of vinclozolin (bottom). The top panel shows that the probability of bias is “probably high” for many papers on many measures, with more than half of papers showing a “definitely high” risk of bias for confidence in the exposure characterization. The bottom panel shows the risk of bias from individual studies.
Risk of bias summary and heatmaps of vinclozolin and radiation animal transgenerational studies. A) Risk of bias bar chart presenting the summary percent ratings for each risk of bias question for the example of animal transgenerational studies. The vinclozolin and radiation exposure studies were used as examples to illustrate internal validity or risk of bias issues for studies of transgenerational design because these exposures were the largest bodies of evidence. B) The risk of bias heatmap of the individual studies of animal vinclozolin exposure.
You can see from these images that much of the risk of bias seems to arise from the failure to report specific aspects of the methods and results. Nine of the fifteen papers listed in this panel are from the lab that performed the study in question. The areas identified as being of high risk for bias are also problematic in the current study as we will go through in detail below. This doesn’t necessarily mean that the studies are flawed or the results are biased, but it does mean that the results cannot be accurately interpreted and it is not possible to determine if they are flawed or valid.
View the other parts of our series on transgenerational epigenetic inheritance:
We live in interesting times. The specters of overpopulation and climate change are constantly in the headlines. The possible threat of global food shortages as a result of increased food demand and climate change-induced crop failures is hovering just over the horizon. And we keep hearing the same mantra: we can’t go on producing and consuming food the way we used to. So how can humanity get out of this fix with the minimum amount of societal upheaval and ecological disaster? If we are to fundamentally alter our food production practices, we must start with a bird’s-eye view of the basic biophysical principles of our current food production system. We may need to begin making food without photosynthesis.
Fundamentals of food
Why not begin by addressing the two most basic questions of food production and consumption – namely, what is food and why do we need to eat it? Let’s start with the answer to the second question, which is simply that food is anything that provides us with two things: (1) metabolic fuel to power our thinking, our movements and our development from a fertilized egg to an adult individual; (2) building blocks to make, repair and maintain the tissues of our bodies.
So whatever food is, it has to satisfy those two criteria. There are three categories of chemical compounds that do just that: carbohydrates, protein and fats. (We also require vitamins and trace minerals for maintaining our bodies but for simplicity’s sake we will focus on these three major macronutrients.) So all types of food are in essence some configuration of carbohydrates, protein and fats with some micronutrients thrown in for good measure.
What is food? A collection of carbohydrates, protein and fats, with assorted micronutrients.
All of these three categories of macronutrients can only be made by living organisms. Granted, some organic precursors – like amino acids, which are the building blocks of protein, have been shown to form spontaneously in nature under specific conditions. (This is probably how life started eons ago.) However, the rates of formation are way too slow to sustain contemporary organisms. As animals, our fundamental problem is that we cannot synthesize carbohydrates, proteins or fats directly from inorganic precursors that occur in the surrounding environment. To be more specific, there are three chemical elements that make up the bulk of these macronutrients – carbon (C), nitrogen (N) and sulfur (S), all of which are inaccessible to us in their inorganic forms such as carbon dioxide (CO2) and molecular nitrogen (N2) in the atmosphere as well as dissolved sulfate ions (SO42–) in soils and seawater.
What that means in practice is that another organism first has to assimilate (“fix”) these three elements by incorporating them into their own biomass as carbohydrates, protein and fats before we then can acquire those macronutrients in turn by eating that organism. Pretty much all the carbon that we ingest was originally fixed by photosynthetic organisms like algae and terrestrial plants.
Up until the early 20th century, all the nitrogen that we derived from our food (nearly all in the form of protein) was originally fixed by specialized bacteria in soils and the oceans. Even with the advent of industrial nitrogen fixation (which I will get back to at the end of this post), we still rely on plants, fungi and various microorganisms to convert inorganic nitrogen compounds like ammonia, nitrate and urea into protein. Sulfur, which we also acquire as protein, can be assimilated in its inorganic forms by most organisms with the exception of animals.
The biophysical limits of food production
Now that we have defined what food is, we can start to explore the current limits of global food production capacity based on agriculture, aquaculture etc. What should be clear at this point is that we do not suffer from a shortage of carbon for food production. Consider that in addition to all the CO2 in the atmosphere (more than 700 billion tons), there are essentially limitless reserves of carbonate-containing rocks in the Earth’s crust. What is limiting are edible forms of carbon i.e. those same carbohydrates, proteins and fats I keep mentioning.
So what ultimately limits our global food production capacity is the rate at which inorganic carbon i.e. CO2, is converted into biomass. (An additional problem is that most biomass on the planet is in the form of cellulose that make up wood, leaves and straw. Although cellulose is a carbohydrate, it is not directly accessible to us because we cannot digest it. That is why we keep livestock – they are essentially walking biocatalysts for the conversion of indigestible plant materials like grass and leaves to things we can eat and drink – meat and dairy.)
Taken together, the theoretical upper limit for current global food production practices is determined by the photosynthetic capacity of the entire planet. The total amount of CO2 that is converted into biomass on land and in the seas every year is known as net primary production (or NPP for short), which has been estimated at 105 billion tons of carbon per year. That might sound like a lot (and it is) but all of those 105 billion tons are not accessible to us for food production. At the moment, the proportion of NPP that has already been appropriated by humanity for food, fuel and fiber has been estimated at 25%. Because of the biophysical constraints of contemporary agriculture (i.e. the need for arable land, sunlight, fresh water and favorable climate conditions), it is believed that this number cannot increase much further.
There are some low-tech options out there on how to get around these constraints. One key factor to consider is that photosynthetic activity is not spread uniformly across the surface of the planet. Instead it is concentrated into patches of high rates of photosynthetic carbon fixation, such as grasslands and forests. At the same time there are huge areas where little or no carbon fixation occurs – polar regions, deserts and nutrient-poor zones of the open ocean. If photosynthetic activity can be increased in an area with low natural photosynthetic carbon fixation, this would effectively increase global NPP.
Green circular fields in the desert resulting from irrigation. Image via Wikimedia Commons.
One way to increase photosynthetic activity would be for example by irrigating arid soils for agricultural production, as is done in the Wadi As-Sirhan Basin in Saudi Arabia. The problem of course is that this requires significant amounts of fresh water, which in the case of Saudi Arabia comes from aquifers that eventually will run dry. Another option is to adopt saltwater-tolerant crops, which would enable the expansion agricultural production to coastal regions where the saline soils prohibit the cultivation of conventional crops. You could take this approach one step further and simply move your crops into the oceans by cultivating macroalgae such as kelp. But what other options are there?
The problem with photosynthesis
Since we more or less take for granted that essentially all carbon in our food comes from photosynthesis, the question is rarely asked whether photosynthesis is the optimal way to fix atmospheric CO2 for food production. Photosynthesis has one major drawback, which is its absolute dependence on light energy. Because of this limitation, conventional crop production can only occur in two dimensions.
Proponents of vertical farming will point out that artificial lighting can bypass this problem and allow for stacking of crops in three dimensions. This is true in theory but in practice you quickly run into the issues of cost and energy conversion efficiency. While sunlight falling on a field of crops is essentially “free”, artificial lighting within vertical farming systems will require electricity from an external source. If that electricity comes from solar panels, we must first consider that these panels typically have an efficiency between 10-20%. Add to that the energy conversion efficiency (also known as radiant or “wall-plug” efficiency) when electrical energy goes into an artificial light source and gets converted into light energy.
On top of that you must consider what proportion of the light coming out of the light source that can actually be harvested by the plant for photosynthesis – a spectrum of light is called photosynthetically active radiation (PAR). Granted, recent developments in light-emitting diode (LED) technology look promising with respect to improvements in both energy conversion efficiency and PAR. However, that still leaves the low efficiency of photosynthesis itself, which tends to end up somewhere in the single digits. To cut a long story short, it would be preferable to have a food production system that is not limited to light energy and thereby would not be confined to two dimensions.
There is another perhaps less obvious drawback with a photosynthesis-dependent food production system, which is that the edible biomass (carbohydrates, protein and fats) is often located within the same physical entity – the crop plant itself, as the light-harvesting apparatus. This means that after annual crops such as wheat, corn or soy have been harvested, the fields in which they were grown are suddenly fixing atmospheric CO2 at much lower rates simply because there are few or no plants left in the field to do so. Remember that the key threshold in global food production is the rate at which CO2 is converted to edible biomass.
An empty field is therefore essentially a waste of photosynthetic potential while we wait for the next batch of crops to appear. (This drawback does not apply to perennial food plants such as fruit trees, which retain their full photosynthetic potential after harvest. Of course, some fruit trees will shed their leaves during winter.) So what we would like in the end is a system that also decouples CO2 fixation from the production of edible biomass. This would allow for continuous CO2 fixation independently of the rate at which edible biomass is harvested.
Breaking the NPP barrier
So to re-cap, in order to circumvent the net primary production barrier, we must come up with a way to convert CO2 to edible biomass (carbohydrates, protein and fats) that (1) does not exclusively depend on light energy, and (2) ideally decouples carbon fixation from the generation of biomass edible biomass. As it turns out, this problem was solved more than 50 years ago during the height of the space race.
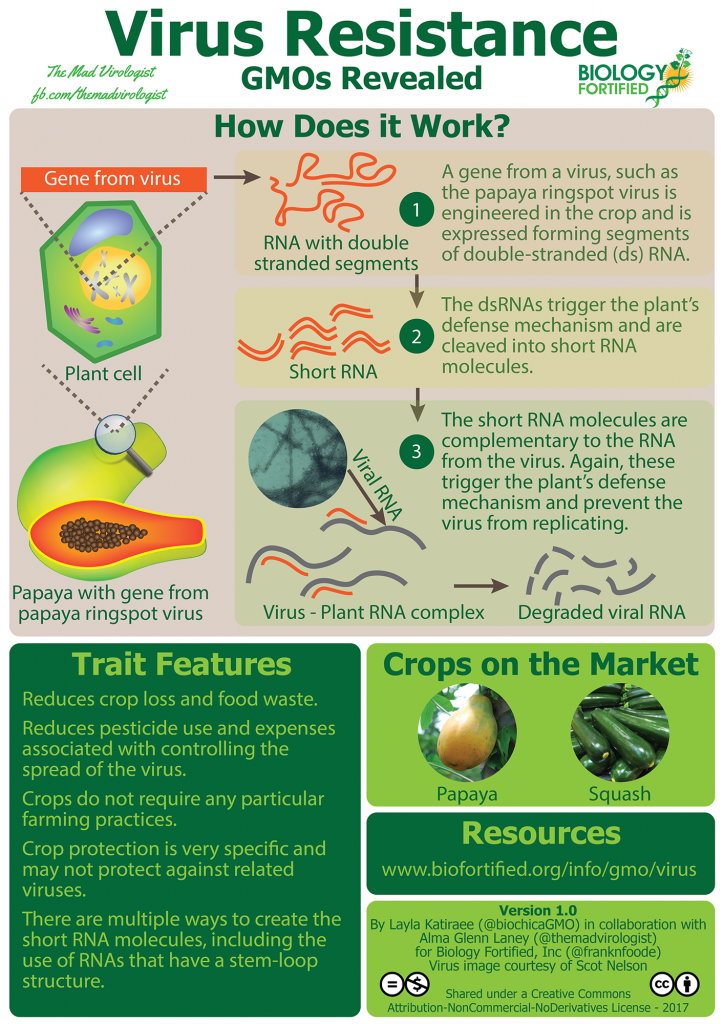
In a 1964 paper, researchers John Foster and John Litchfield described a continuous CO2-recycling life-support system intended for extended space travel, which would capture the CO2 exhaled by astronauts and convert it into food. This was made possible by an edible bacterium called Cupriavidus necator – also known under a host of older names including Ralstonia eutropha. This bacterium is capable of chemosynthesis, which is a process analogous to photosynthesis but relies on chemical energy carriers instead of light to power CO2 fixation. In the case of the C. necator bacterium, it uses the inherent energy in hydrogen gas (H2) to power the CO2 fixation process.
Hydrogen gas can be generated by electrolysis of water, which is the key to satisfying the first criterion of a non-photosynthetic food production system, as the electrical energy can be provided by any energy source – hydro, wind, geothermal and even nuclear. There might even be a point of using solar power for chemosynthetic production, since it would be possible to place solar panels in a place where conventional agriculture is not possible (like a desert) and then transmit the electrical energy through the power grid to wherever the hydrogen production and subsequent cultivation of edible bacteria takes place. This property also partially satisfies the second criterion of a non-photosynthetic food production system by decoupling one aspect of CO2 fixation (generation of energy) from production of edible biomass. The enzymatic machinery inside the C. necator cell is still required to convert CO2 into biomass. However, this is not a major problem since a C. necator cell growing on a H2/CO2 mixture will divide fairly quickly under optimal conditions – as often as once every three hours. This means that it is possible to harvest half of the C. necator bacteria every three hours indefinitely as long as the bacteria are continuously supplied with a H2/CO2 mixture.
Solein is an example of a protein produced without photosynthesis. Image used with permission from Solar Foods.
There are in fact some companies today that are trying to commercialize food and feed products derived from H2-dependent chemosynthetic bacteria like C. necator. The Bay Area startup NovoNutrients use industrial CO2 emissions to cultivate an undisclosed microorganism – most likely a H2-dependent chemosynthetic bacterium, which is then processed into a high-protein flour called Novomeal, which is then marketed as aquaculture feed. The Finnish startup Solar Foods are working on a similar process but intends to market the resulting protein product – called Solein, for human consumption rather than be used as animal feed. Solar Foods recently announced that they plan to have Solein for sale in supermarkets by 2021.
The idea of eating protein flour made from bacteria might seem like a novel idea but the concept of edible microorganisms such as bacteria, yeasts and filamentous fungi is in fact quite old. The Aztecs were known to harvest photosynthetic bacteria from Lake Texcoco before the lake was drained following the Spanish conquest (the remaining lake basin is now part of Mexico City). The same kind of photosynthetic bacteria are still harvested today from alkaline lakes in Chad by the indigenous Kanembu people (as described in the video below). In 1902 the Marmite Food Company in the UK launched its now (in)famous sandwich spread made from spent brewer’s yeast. More recently, biomass from the filamentous fungus Fusarium venenatum is used as the main ingredient in Quorn-brand meat imitation products.
Microbial biomass makes a good source of food due to its high protein and vitamin content. Unfortunately, only a small minority of edible microorganisms is capable of chemosynthetic growth like that of C. necator. Most edible microorganisms instead require organic substrates for growth – sugar being the most commonly used substrate for microbial biomass production at present. However, most microorganisms also have the ability to grow using very simple organic compounds like hydrocarbons, alcohols and organic acids.
As it happens, there are a number of hydrocarbons, alcohols and organic acids that can be synthesized directly from CO2 using different chemical and biological processes. This means that rather than edible microorganisms fixing CO2 themselves, the CO2 is fixed in a separate process to produce an organic compound that the edible microorganism can then use for growth. Such a two-step process would satisfy the two criteria for non-photosynthetic food production mentioned previously, namely (1) not being solely reliant on light energy and (2) decoupling carbon fixation from generation of edible biomass.
There are several possible options for how edible microbial biomass could be produced from organic compounds that in turn have been synthesized directly from CO2. I will describe one such example in more detail. Methanol (CH3OH) is the simplest alcohol and can be produced from CO2 by a simple hydrogenation reaction:
CO2 + 3 H2 → CH3OH + H2O
The company Carbon Recycling International runs a factory on Iceland, which produces 4,000 tons of methanol per year by hydrogenating CO2 from emissions that come from a neighboring geothermal power plant. This methanol is intended for use as a drop-in fuel but could also be used to cultivate a wide variety of edible microorganisms. Although methanol is toxic to animals, many microorganisms can grow just fine using methanol as their only source of metabolic carbon. In fact, during the 1970s and 80s, the British company Imperial Chemical Industries developed a high-protein animal feed called Pruteen, which was derived from the edible methanol-assimilating bacterium Methylophilus methylotrophus.
Ultimately Pruteen production was discontinued due to rising methanol prices and competition from cheaper soy-based feeds. Nevertheless, the Pruteen process demonstrated that it was possible to produce edible microbial biomass in significant amounts (50,000-60,000 tons per year) from a single 1,500-m3 bioreactor. I have recently estimated that if the entire US soy production capacity (about 120 million tons per year) was to be replaced with methanol factories and Pruteen-style bioreactors, it would occupy roughly one thousandth of the same land area. However, this estimate does not include energy sources to power the CO2 conversion process. So the land-sparing potential of such a process would depend heavily on the power density of the chosen energy source.
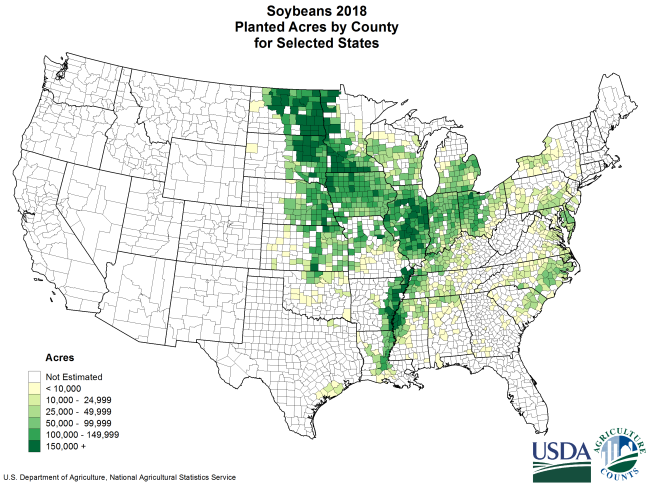
In 2018, 89.6 million (89,600,000) acres of soybeans were planted in the United States in 2018. If bioreactors were used instead, only 89.6 thousand (89,600) acres would be needed to produce the same amount of protein.
Final big thoughts
What should be clear from the examples of edible chemosynthetic bacteria as well as edible methanol-assimilating bacteria that I presented above is that producing edible microbial biomass from CO2 in a manner that does not require photosynthesis carries with it some pretty significant implications. Microorganisms are typically cultivated in large (10-1000 m3) bioreactors where internal growth conditions (temperature, rate of mixing, supply of air and nutrients etc) are controlled independently of external conditions. This means that edible biomass – food, can be produced anywhere the planet independently of local climate conditions or access to arable land. Suddenly deserts, tundra, underground caves or even the open ocean can become hubs for high-capacity food production.
If global food production capacity is no longer constrained by NPP, the new limits for just how much food humanity can produce per unit time now comes down to energy and money. What will this mean for Earth’s carrying capacity? Will the human population continue to grow if food production is no longer constrained by photosynthesis (and prove Paul Ehrlich wrongagain)? If my back-of-the-envelope calculation is correct, will the higher production density of edible microbial biomass enable us to restore a significant proportion of agricultural land to its natural state thereby protecting biodiversity – and perhaps even managing to sequester a big chunk of atmospheric CO2 in the process? It all remains to be seen.
In closing, it might be instructive to compare our current situation with the scarcity of nitrogen fertilizers (manure, nitrate-containing minerals) that faced global agriculture at the end of the 19th century. At the time it seemed as if the global food production system had reached a seemingly insurmountable boundary. Then came the invention of industrial nitrogen fixation by German scientists Fritz Haber and Carl Bosch in the beginning of the 20th century and suddenly humanity was no longer dependent on biological nitrogen fixation. As a result the human population could continue to grow from less than two billion in the year 1900 to 7.7 billion today (and counting). In fact it has been estimated that half of the nitrogen in our bodies now comes from the Haber-Bosch process rather than biological nitrogen fixation.
If humanity can become independent of biological carbon fixation as we did with nitrogen, recent history would suggest that this in itself will not promote stabilization of the global population. Instead the global population is expected to plateau and eventually shrink thanks to falling fertility rates as a consequence of increasing living standards, which allow parents across the globe to choose to have fewer children. So ultimately the challenge of this century (and perhaps the next one as well) will be to feed humanity in a sustainable fashion as it attempts to clear this demographic hurdle. And if you ask me, edible microbes produced independently of photosynthesis would seem like the way to go.
Written by Guest Expert
Tomas (“Tom”) Linder is a microbiologist and molecular geneticist who studies metabolism in microorganisms and has a particular fondness for yeasts. He is based at the Swedish University of Agricultural Sciences in Uppsala, Sweden. Follow Tom on Facebook at Yeast Genomix.
Developers of the fast-growing genetically engineered salmon first started the approval process with the United States (US) Food and Drug Administration (FDA) in 1995. In 2019, we are a bit closer to having the option of buying this fish for ourselves, as the FDA clears the final regulatory hurdle to allow sale of AquAdvantage salmon.
The resulting genetically engineered fish are ~99.99986% Atlantic salmon, with the addition of just 4,205 base pairs in a genome of 2.97 billion bases. Further, the growth hormone proteins from Chinook and Atlantic salmon are 95% identical. This leaves the ocean pout promoter as the only “new” element. The developers chose this promoter because genes it controls are continually expressing – always on – as opposed to the salmon promoter for growth hormone, which is only on in certain environmental conditions.
Even though the inserted growth hormone gene is always on, it doesn’t have much of an effect unless the fish have access to food. When allowed to eat as much as they want (fed to satiation), juvenile AquAdvantage salmon can grow nearly 3 times longer than conventional juvenile Atlantic salmon. But in a simulated natural environment with limited food, juvenile AquAdvantage salmon grew only a little larger than juvenile conventional salmon.
While AquAdvantage salmon grow faster, they do not grow larger overall – adult AquAdvantage salmon and adult conventional Atlantic salmon are the same size. The genetically engineered fish just get to that size faster and with less feed. As FDA describes, “the overall total amount of feed required to produce the same fish biomass was reduced by 25%” for AquAdvantage salmon.
There will always be a place for wild-caught fish on our plates, just as there is a place for other speciality products like heirloom pork, wild mushrooms, and so many other wonderful things. But in a world with a growing population and a growing demand for nutritious fish like salmon, we can not rely only on wild stocks. Aquaculture is a necessity.
Wild-caught Pacific salmon are not able to meet demand, in part due to vulnerability to higher water temperatures related to climate change. In 2018, US west coast salmon fisheries along Washington, Oregon, and California requested fishery disaster assistance from the Department of Commerce due to commercial fishery failures in 2015, 2016, and 2017.
Even as wild fisheries decline, demand for salmon is increasing. In 2016, the US imported ~339,000 metric tons of salmon, mostly farmed Atlantic salmon raised in cages in the ocean, as Richard Martin reports in Genetically engineered fish is not a matter of “if” but “when”. In 2018, the US imported 403,107 metric tons of all types of salmon, with a value of over $4.1 billion, (327,116 metric tons and $3.4 billion of Atlantic salmon alone) according to the National Marine Fisheries Service.
The Monterey Bay Aquarium Seafood Watch rates indoor farmed salmon (raised the way genetically engineered salmon would be raised) as a “Best Choice.” Fish that grow faster will consume less feed, take less energy, decrease prices for consumers, and potentially provide a product with fewer parasites than ocean-raised fish. All this, with the potential to create jobs in the US with additional rearing facility locations, and to decrease reliance on ocean-based farms and open-air ponds that pose known risks to ecosystems.
In the US, AquAdvantage salmon required two assessments: one to determine safety of a new animal drug that would be entering the food supply (under the Coordinated Framework for Regulation of Biotechnology, new genes and gene products in genetically engineered animals are regulated as animal drugs by the FDA), and one for potential risk to the environment (as required under the National Environmental Policy Act).
In Canada, AquAdvantage salmon required three assessments: one for safety and nutrition of the salmon for use as food, one for safety and nutrition of the salmon for use as a livestock feed, and one for potential risk to the environment.
The regulatory process may have been lengthy, but one benefit is that we can be confident that the salmon is safe to eat and safe for the environment. We have two independent sets of regulatory processes to consider, one in Canada and one in the United States. While there are similarities in what the regulatory agencies are looking for, the laws and regulations were developed independently and the people in the agencies themselves are different, under different types of internal and external influences.
The agencies in both the United States and Canada independently found fast-growing genetically engineered salmon to be safe to eat and safe for the environment. A summary of the safety findings from the regulatory agencies is below.
GMO salmon for food and feed
The regulators considered three main types of issues with regard to using AquAdvantage for food and feed: nutrients, hormones, and allergenicity.
Protein, fats, vitamins, and other components of fish and other foods can vary widely due to the variety of the plant or animal, the place it was raised, the weather while it was growing, and many other factors. Because there is so much variation within each food type, a simple comparison between genetically engineered and non-genetically engineered doesn’t make sense. Instead, scientists and regulators look at a range of values.
For example, a Consensus Document for maize reports that protein in field corn ranges from 6% to 12.7% of dry weight. Therefore, in testing the protein of corn with genetically engineered traits, we’d need to take a second look at any values that fall outside of that range.
Fast-growing genetically engineered salmon could make nutritious and delicious dishes like this more sustainable and affordable.
Nutrients in GMO salmon
Both FDA and Health Canada determined that fast-growing genetically engineered salmon is safe for humans and animals to eat. Both agencies found that there were no substantial changes in the protein and fat (including omega-3 fatty acids) profiles of AquAdvantage salmon compared to expected ranges.
For diploid AquAdvantage salmon, the niacin level was higher than expected ranges, but the level is not of nutritional concern. The majority of AquAdvantage salmon salmon (more than 99%) are triploid, and triploid salmon had niacin levels within expected ranges.
Hormones in GMO salmon
Specific concerns with AquAdvantage salmon include increased hormone content in the edible parts of the fish. Both FDA and Health Canada concluded that there is no health risk to humans or livestock associated with hormones in AquAdvantage salmon.
First, the growth hormone produced by AquAdvantage salmon is from Chinook salmon. Chinook salmon are commonly consumed and have a safe history of use, with no concerns about toxicity. Second, the amount of growth hormone (and associated regulatory hormones) in muscle and skin of adult (market weight) AquAdvantage salmon was not distinguishable from conventional salmon. People who eat AquAdvantage salmon would not be exposed to any more growth hormone than people who eat conventional salmon or other animal-sourced foods.
Health Canada points out that “the scientific literature has consistently shown that dietary growth hormones from various animal species have very poor bioavailability in mammals and humans.” This means that growth hormones consumed in the diets are primarily digested, not absorbed intact. Any tiny amount of the Chinook salmon growth hormone that might be absorbed would have a limited effect in humans “due to the species specificity exhibited by the human [growth hormone] receptor.”
Allergens in GMO salmon
Fish, including salmon, is a major food allergen. Of course, people who are allergic to salmon will also be allergic to AquAdvantage salmon. Still, the regulatory agencies investigated whether AquAdvantage salmon would have more fish allergens than conventional salmon. Analysis showed that allergens in triploid AquAdvantage salmon were within the normal range found in conventional salmon.
Diploid AquAdvantage salmon (less than 1% of AquAdvantage salmon) did have statistically higher levels of allergen content, but experts concluded that there would be no impact on non-allergic consumers, and no change in allergenicity for allergic consumers.
GMO salmon in the environment
If reproductive-capable, fast-growing salmon were to escape captivity, they could theoretically breed with nearby salmon or trout and spread the gene for fast-growth. This could have negative consequences for the salmon population, their food sources, competing fish species, and other parts of the ecosystem.
When AquaBounty requested FDA approval of their AquAdvantage salmon, they were very specific about how and where the fish would be raised. The request was for one egg production facility in Prince Edward Island, Canada and one fish production facility in Panama. AquaBounty later requested that a fish production facility in Indiana be added. FDA’s approval is for these locations only, and additional approval would be needed for more locations.
AquaBounty selected (and FDA approved) these locations to have many overlapping ways to prevent release of GMO salmon into the environment. The containment methods include biological, physical, and environmental aspects.
AquAdvantage salmon will not be raised in an environment where they can exhibit this leaping behavior.
Biological containment
Part of biological containment is Atlantic salmon’s own biology. Atlantic salmon reproduction requires fresh running water over a gravel bed. Salmon have complex mating and nesting behavior, and a sexually compatible male must be present when the female spawns. In the waters near the egg and fish rearing facilities, there are no males or gravel beds. Even if a female were to spawn, the eggs would not be fertilized. In addition, spawning takes so much energy that 60% or more female salmon die after spawning.
Another part of biological containment is that AquAdvantage salmon can not breed – they are sterile. AquaBounty pressure treats salmon eggs to induce triploidy (3 sets of chromosomes instead of 2 sets). Since a small percentage of triploid males can reproduce, AquaBounty adds another layer of biological containment by only using female fish. A 2018 study, Comparisons of reproductive function and fatty acid fillet quality between triploid and diploid farm Atlantic salmon, confirmed that “escaped triploid farm salmon are very unlikely to reproduce in the wild.”
If you eat fish, you’ve likely already dined on triploid fish. Triploid fish of many species, including salmon, trout, and carp, have been used around the world for decades. They are used in commercial fisheries and recreational fishing areas to prevent farmed or stocked fish from breeding with wild fish. Triploid fish grow to a larger body size and have higher quality meat because the animals do not undergo the stress of reproduction. Other triploids that we eat include bananas and watermelon.
Lastly, the fast-growth trait serves as a sort of biological containment as well. Fast-growing fish (whether resulting through breeding or biotechnology) and triploid fish have some changes relative to conventional diploid salmon that could decrease survival in the wild. Changes that may affect AquAdvantage salmon survival in the wild include: increased metabolism causing smaller energy reserves, higher oxygen consumption, decreased tolerance to stress, increased appetite, increased aggression, and reduced response to predators.
Physical containment
Because a small percentage of AquAdvantage salmon could be capable of reproduction, additional containment methods are necessary to ensure eggs or fish do not escape. Their facilities use numerous layers of filters, screens, and nets. Chlorine is used in drains to kill eggs or fry that might somehow slip through. Facilities are inspected daily and standard operating procedures are in place for every process in each facility.
AquaBounty also has security to protect against human sabotage. Facilities have security cameras (the Panama facility has guard dogs) and fencing around each property (fencing is topped with barbed wire at the Panama and Indiana locations) among other measures.
Security cameras are part of the physical security measures used by AquaBounty at their facilities.
Environmental containment
In the highly unlikely case that a fertile AquAdvantage salmon could escape, the local environment near each facility would make it nearly impossible for any escapee to survive to adulthood, find a sexually compatible male fish, and reproduce.
The egg production facility is located in Prince Edward Island, Canada. The biological and physical containment measures mean it is nearly impossible that a fertile egg would make it past the filters and chlorine. Any that did would not find a good place to grow into adult fish. The eggs are raised in fresh water, so the salinity in the nearby river reduces likelihood of survival. In the winter, water near the facility is too cold for salmon. Any escaped eggs that somehow managed to grow into adult female fish would not find a male to mate with. Atlantic salmon previously lived in this area, but overfishing, barriers to migration, and acid rain have made them locally extinct.
The fish production facility in Panama is located at a high altitude near a river that drains to the Pacific ocean. Much of the river water is used for power generation, and canals that control water flow are not suitable for salmon. Dams provide a physical barrier to movement downstream. If any fish escaped and they managed to get past the barriers, they could potentially survive in the river closest to the facility, but they would not find any males to mate with. If they tried to move out to sea, high temperatures in the lower lower parts of the river would kill the salmon.
The fish production facility in Indiana includes a series of ponds that will be seeded with wetland plants to help clean effluent before it drains away from the facility. Effluent from the ponds must pass through a screen before reaching a drainage ditch that ultimately drains into the Mississinewa River. The ditch is dry except during wet weather. Even with the effluent from the facility, the water levels in the ditch are expected to be only 1-2 inches near the facility. If fish somehow managed to get out of the facility, past the ponds with screen, and past the ditch into the Upper Mississinewa watershed, they may find conditions where they could survive during some times of the year. However, the watershed is too warm with too little oxygen and too much dissolved solids for salmon and related species like trout. A survey of the watershed found no species of trout or other cold-water salmonids at any of the 35 sampling sites, so any escaped females would not find a male with which to mate. There is also a dam downstream of the facility that would prevent movement of fish, if they were to somehow survive.
The FDA states that “at full capacity, the [Indiana] facility will harvest approximately [90 metric tons] of AquAdvantage Salmon per month.” This will meet only ~0.27% of US demand for salmon (403,107 metric tons in 2018). If there is reasonable consumer acceptance of AquAdvantage salmon, AquaBounty may request approval for more facilities from the FDA. Those facilities will be regulated and inspected by FDA and appropriate agencies in the state in which they are located, continuing to keep appropriate environmental containment of the fast-growing genetically engineered salmon.
Italian researchers Elisa Pellegrino, Stefano Bedini, Marco Nuti & Laura Ercoli published a meta-analysis of the impacts of genetically engineered (GE) corn on the environment, agriculture, and toxicity. The fully study is publicly available: Impact of genetically engineered maize on agronomic, environmental and toxicological traits: a meta-analysis of 21 years of field data. I encourage you to read the study itself, and I provide a summary with commentary below.
A meta-analysis is a formal way to combine many papers on related topics to come to some overall conclusions (see 5 key things to know about meta-analysis). There have been large GE crops literature reviews in the past, but none to date have actually compiled all of that data into a meta-analysis.
The researchers reviewed 6,006 studies in the peer-reviewed literature from 1996 to 2016, including papers on yield, grain quality, target organisms (pests), non-target organisms, and soil biomass decomposition. Amusingly but not surprisingly, most of the studies took place in the US corn belt: Iowa, Illinois, and Nebraska. The researchers excluded experiments that were not performed under field conditions, that did not use a genetically similar (near isogenic) comparator, that had GE and non-GE corn grown under different conditions, or that had small sample sizes or other statistical issues. That left only 76 publications in the meta-analysis.
One drawback to this meta-analysis is that it groups GE traits in corn together. Each type of GE trait has benefits and drawbacks and typically must be considered individually. That said, in public discourse, specific GE traits are rarely discussed and people simply say “GMOs”. In that context, this meta-analysis is helpful in answering concerns about the effects of “GMOs”. The researchers “noted that single event [herbicide tolerant] hybrids were missing”. In other words, studies included GE herbicide tolerance only when combined (stacked) with GE insect resistance traits. Effectively, this is a meta-analysis on corn with GE insect resistance traits, not on all currently available GE corn. Modified figure 3a from “Impact of genetically engineered maize on agronomic, environmental and toxicological traits: a meta-analysis of 21 years of field data”.
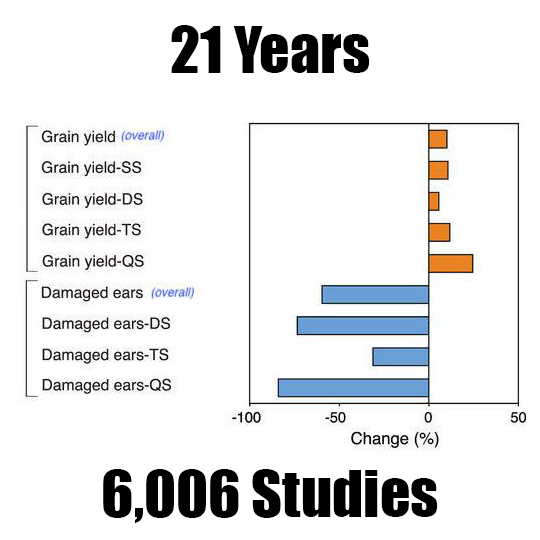
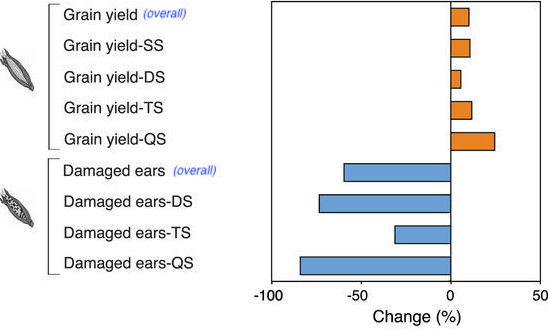
The results of the meta-analysis are striking. As you can see in Figure 3a from the paper, corn yield wass increased overall in GE corn compared to non-GE corn. There were small differences in how much yield was increased depending on whether the varieties had 1, 2, 3, or 4 biotech traits (single stack, SS; double stacked, DS; triple stacked, TS; and quadruple stacked, QS), with quadruple stack having the highest yield.
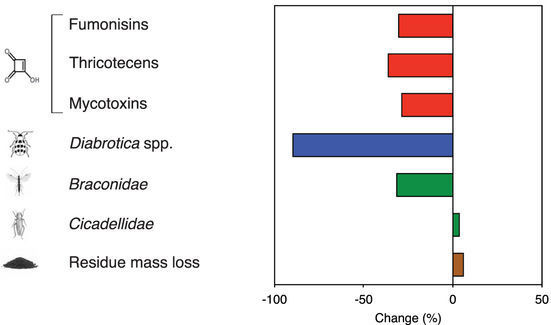
There were also fewer damaged ears overall in GE corn compared to non-GE corn. This is important because damage to the corn leads to fungal infection, which leads to potentially deadly mycotoxins. There was no significant difference in ear damage between single trait varieties and genetically similar non-GE varieties, but there was a significant difference in yield, which is interesting because you’d expect these two measurements to be more correlated. It’s also interesting that quadruple stacks had much less damage than triple stacks, but only a little less than double stacks. Figure 3b from “Impact of genetically engineered maize on agronomic, environmental and toxicological traits: a meta-analysis of 21 years of field data”. Figure 3b shows that overall GE corn has less mycotoxins, great news since mycotoxins can cause all sorts of ill effects in humans and animals who eat them. The rate of decomposition (residue mass loss) was higher in GE corn than in non-GE corn.
The major pest corn rootworm (Diabrotica) was reduced in GE corn, not a surprise since that is the target pest of many GE insect resistance traits. Beneficial parasitic wasps (Braconidae) were also decreased in GE corn, possibly due to greatly reduced numbers of corn rootworm hosts. The minor pest leafhoppers (Cicadellidae) were increased in GE corn, possibly because the use of insect resistance traits reduces the need for broad spectrum insecticides.
Many beneficial arthropods were not affected by the use of GE corn: pirate bug (Anthocoridae), spider (Araneae), ground beetle (Carabidae), lacewings (Chrysopidae), lady bug (Coccinellidae), damsel bug (Nabidae), sap beetle (Nitidulidae), rove beetle (Staphylinidae). Pest aphids (Aphididae) were also not affected by the use of GE corn. This was surprising. I expected that there would be higher prevalence of many beneficial and pest insects in GE corn compared to non-GE corn due to reduced treatments with broad spectrum insecticides.
Another not-very surprising finding was that there were no significant differences in grain quality: proteins, lipids (fats), or fiber. There was also no difference in the lignin in stalks or leaves. This confirms that corn is corn, whether it is GE or not. It would be nice to see meta-analysis on minor constituents of GE and non-GE corn, such as particular vitamins or limiting amino acids. The researchers for this study add biodiversity and soil biogeochemical cycles to the wish list of items they’d like to see studied. Even with the limitations of what is available in the literature, this meta-analysis shows once again that crops produced with biotechnology are some of if not the most studied foods that we eat.
The AquAdvantage Salmon is engineered to grow faster and could be fed Omega-3 enhanced crops like camelina.
Biotechnology in the fish farming industry hit the headlines in 2017 when a fast-growing salmon became the first GM animal to reach the market. In Europe, however, new developments in biotech’s potential for aquaculture went largely un-reported. Rather than focusing on the fish, British scientists are making progress towards creating fish food from a transgenic plant. For almost two decades, scientists from Rothamsted Research and their collaborators have been working towards providing oils for farmed fish from one of Europe’s oldest oilseed cropsLcamelina. The latest results bring them a step closer to realizing their vision.
Why focus on fish feed? To understand that we must think about where our fish and their nutrients come from.
The current problems with meeting our demand for fish
Over half of the fish we consume comes from farms, and aquaculture uses around 80% of the fish oil harvested annually from the sea. More fish goes into the system than comes out, which means that much of the fish caught from the wild goes to feeding farmed fish. Growing demand for omega-3 LC-PUFAs, particularly from aquaculture, has placed substantial pressure on the wild fisheries which supply much of this fish oil. Camelina seeds. Credit: Rothamsted Research
The root of the problem is that that fish oils aren’t produced by fish. Instead fish acquire them from microalgae, which aren’t available to caged fish in farms. We don’t currently have financially-viable technology to produce algae on a large scale, so farmed fish are fed oil and meal made from fish caught from the oceans. In this way, the fatty acids consumed by the smallest fish accumulate all the way up through the food web.
Vegetable oil can be used as an alternative to fish oil in feed, but this changes the nutritional value of the fish people eat. Given that fish can’t make their own fish oils, they can only accumulate these oils if they consume them as part of their diet. Now that farmed fish have more vegetable oil in their diets, they are no longer providing consumers with the same level of omega-3 and the associated health benefits.
Producing the appropriate oils from plants has the potential to ensure that farmed fish bring their full benefits for human health, whilst simultaneously reducing pressure on wild fish stocks.
Promising results from 2017
Fatty acid profiles of wild-type and GM camelina seeds in both the lab and the field. From Usher et al. 2017
A new paper in Scientific Reports shows that their plants have the potential to yield oils in the field, and results published in the journal PLOS ONE showed that these oils are suitable food for salmon. The project, led by Professor Johnathan Napier, uses Camelina sativa (false flax), a distant relative of oilseed rape. Camelina is naturally high in short-chain omega-3, but it’s long-chain omega-3 that is important for fish food. To change the profile of oils in the seed, the team has introduced synthetic DNA sequences into the camelina genome. The introduced genes are similar to those found in marine microbes, and code for enzymes in the biochemical pathway which produces long-chain omega-3 polyunsaturated fatty acids (LC-PUFAs).
The oil can be extracted from the seed and fed to fish, and that is exactly what happened in the feeding trial reported in PLOS ONE. The newly-published data show results from a 2015 field trial of GM camelina, and confirm positive findings from 2014.
Firstly, results from both years show that the transgenic plants perform well in the field. The studies went beyond agronomic performance, as they also examined the composition, or profile, of the oil produced by the plant.
The profile of seed oil has also been improved relative to earlier varieties of transgenic camelina. Not only were the scientists looking to increase the concentration of desirable fatty acids, they were also keen to reduce the production of some of camelina’s natural oils. Omega-6 fatty acids are only present in low levels in marine organisms, so the latest varieties have been engineered to produce smaller quantities.
Salmon feeding trials have also seen positive results, and oil from GM camelina was an effective substitute for fish oil in feeds, supporting growth without compromising fish health. The fish accumulated fatty acids exactly as they would when fed oil from fish, creating a fish with the desired health benefits for consumers.
Trials on sea bream have also indicated that GM-derived oils could replace fish oils in their diets.
The future of camelina
The results published in 2017 indicate that oils from transgenic plants offer the opportunity to increase the levels of omega-3 LC-PUFAs in farmed fish to levels found a decade ago. Further development work is now taking place to make this a reality on a large scale.
The next steps for the project is to select the best camelina lines – i.e. those producing the highest levels of omega-3 fish oils. These plants will be used in larger field trials to demonstrate that they perform as well in the field as they do in the glasshouse. Cross-sections of camelina seeds showing increased levels of Omega-3 fatty acids. From Usher et al. 2017
The work will then need to move from the research phase to development and commercialization. As well as planning how to make the crop available on a commercial scale, an essential step will be seeking regulatory approval. Although the research has been done in the UK, the status of the EU regulatory system means that the team will be applying for approvals outside of Europe.
Issues to be addressed before the plants are suitable for regulatory approval include freedom-to-operate (ensuring that no patents would be infringed if the crop is commercialized), and appropriate farm management. Camelina is predominantly self-pollinated but is visited by insects, so during the trials scientists addressed concerns from local beekeepers by covering the flowering crop with a fine mesh net to prevent bees from transporting GM pollen back to their hives. Such measures wouldn’t be feasible if the crop was grown on a commercial scale, but the team is considering the farm management practices which would be needed to prevent gene flow. Thankfully, camelina doesn’t cross pollinate with oilseed rape or other Brassica species, making large-scale planting more feasible without risking cross-pollination of other crops. In the end, to goal is to make aquaculture more environmentally sustainable while maintaining the healthy properties of the fish that we eat.
For updates on the project and more background reading, please visit the Rothamsted website. I also cover the topic in my book Is that Fish in Your Tomato?
Written by Guest Expert
Rebecca Nesbit is author of the popular science book ‘Is that Fish in your Tomato?’ which explores the fact and fiction of GM foods. She studied butterfly migration for her PhD, then worked for a start-up company training honeybees to detect explosives. She now works in science communication and her projects have ranged from a citizen science flying ant survey to visiting universities around the world with Nobel Laureates. In her spare time she writes fiction – she has published a novel, and many short stories.
Last year while driving across the country, I made a slight detour northward to visit the city of Moscow, Idaho. I met up with Professor Shelley McGuire at Washington State University and two other members of her team, Kimberly Lackey and Bill Price, who together published the study that showed that breast milk did not contain glyphosate, the active ingredient in Roundup. It was a simple and scientifically straight-forward series of experiments, and was confirmed by a separate research team in Germany. Scientifically this was uncontroversial, but Dr. McGuire’s team became embroiled in controversy because some organizations found it inconvenient for their political campaigns against the chemical. The threatening postcard I received, similar to the one mailed to Shelley McGuire.
Organizations like Moms Across America and Sustainable Pulse attacked her reputation and her research, while USRTK submitted records requests for all her correspondence. This study was so important that they gave her both barrels. Meanwhile, she was open, patient, and wanted to answer people’s questions. Followers of these organizations even got upset when their moderators deleted her civil answers to their questions. It became apparent that they didn’t even want their followers to learn from this study.
At the height of it all, Shelley received a harassing postcard. With a combination of bad photoshop skills and maximum creepiness, someone sought to harass her by snail mail. We know of only one other scientist who has received such an illuminating treatise on productive dialog – me*. The Boston postal artisan even wrote “Thanks for the support” in German, giving it an original charm it desperately needed. When people lash out at scientists with such hateful, tasteless, confused artifacts, it means that they are revealing parts of the universe that are dangerous to their identity.
In Moscow, we sat down and had a great chat about the study, its methods, interpretations, funding, and public reactions. I also described my visit to Covance in Madison, WI, which is the company that did some of the glyphosate detection experiments. I learned that conflicts of interest are not always what they seem. Dr. McGuire’s research was thorough, confirmed, and influential for public policy – everything that the organizations who attacked her were not. This contrast was also explained in Food Evolution (see my review here), which you should see if you haven’t yet done so.
In all these serious considerations of glyphosate, breast milk, science and conflict, we found some good opportunities to laugh. Give it a watch, and tell us what you think! After all, no subject is too heavy to grab a refreshing drink, sink into a comfy couch and have a nice evening chat.
Reference:
McGuire, MK et al. Glyphosate and aminomethylphosphonic acid are not detectable in human milk. Am J Clin Nutr. 2016 May;103(5):1285-90. doi: 10.3945/ajcn.115.126854 http://ajcn.nutrition.org/content/early/2016/03/30/ajcn.115.126854.abstract
“Our data provide evidence that glyphosate and AMPA are not detectable in milk produced by women living in this region of the US Pacific Northwest. By extension, our results therefore suggest that dietary glyphosate exposure is not a health concern for breastfed infants.” *After publishing this article, several other individuals have come forward and reported to have also received a version of the skull-and-crossbones postcard. Mary Mangan published a Storify cataloguing them as they come forward.
I’m excited to announce today that our first Citizen Science experiment, the “GMO Corn Experiment” is entering its final stage. It has been a long time coming, and I know each and every one of our participants have been waiting to hear the news about our results. I have a lot of things to tell you about in this update, and I’m happy to say that while we encountered some issues with analyzing all the voluminous data that our citizen scientists contributed, we have our results and are preparing them for publication. Along with this update, we want to make sure that every one of our citizen scientists gets a chance to get credit in the final paper.
I’m going to tell you a story spanning the beginning of the experiment right up to today. So to make sure everyone hears the news and gets the takeaways, here is where we are. The analysis of the data from the GMO Corn Experiment is complete, and we are preparing our study for publication. We encountered many difficulties when it came to analyzing all the data submitted to the experiment, but we worked through them and have some clear results that we presented at a scientific conference to get feedback from fellow scientists. We have verified the genetic identity of the ears of corn used in the experiment, and confirmed that they are equivalent in composition. We are sending out a survey to our Citizen Scientists so we can credit them in the paper, and when the paper is submitted we will hold a live broadcast announcement and release some of the results and award prizes to some of our participants who have completed our survey by August 25. Want to know all the details? Read on!
On data and difficulties
Several years ago, when I initially envisioned this project, I thought it would be nice to have 30, 50, or maybe even 100 experiments. That would give us some good numbers to get a satisfying conclusion. As we prepared to announce the project in late 2015, we were going with that plan. Long before the public announcement of the experiment, I requested ears of corn from Monsanto that we could use to test the claim that wild animals would avoid genetically engineered corn, and they agreed to grow two plots of corn in Hawaii and ship the ears to us. As we prepared for that public announcement, I got an email from Monsanto saying that they harvested 3,000 ears from each plot, and wanted to know how many I wanted. Hmm, how about all of them? 100 experiments turned into 2,000 potential experiments in 1,000 kits, thanks to our enormously successful fundraiser. If some data is good, more is better, right?
A year later, we had hundreds of completed experiments, and about 2,000 observations for us to go through. (Well, for me to go through!) As exciting as it was, this was a daunting task. When planning the experiment, we considered all kinds of ways that our citizen scientists could enter their data, and decided that sending photos would be the best way since we could always refer back to them if there was any confusion.
But analyzing images presented its own problems: how do we make sure that the numbers would be reliable and repeatable? The first thing we tried was to analyze the images based on color. The yellow maize kernels have a specific color, and the images can be analyzed to count up the pixels showing how much of the corn is left on either side of the image. By converting the images into false colors it would be just a matter of adding up pixels. Not only would it be repeatable but it would be done entirely without human judgements and whatever biases might be there. Reliable.
We knew that not all of the images would fit this kind of analysis. Some were blurry or taken at a distance, but if enough experiments made the cut we could use those to make our conclusions. It failed. There was simply too much variation in angles, distances, camera types and more to get good data out of this, as Kevin explained in an update he included in his podcast, Talking Biotech. It would be an easy thing to do if this experiment was done in a lab with a solid black background and a precisely positioned camera, but out in the wild as it were, we needed to find a better way.
Settling the score
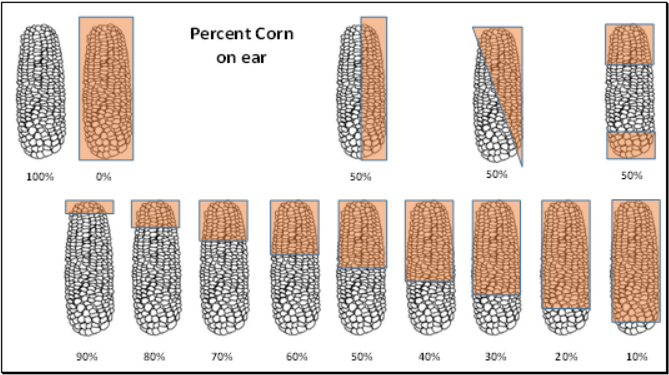
As plant scientists, we went back to our roots with phenotyping. Plant biologists walk through fields rating plant phenotypes on number scales, so we applied that approach to the images. Anastasia Bodnar created a scale, and Kevin Folta gathered a group of volunteers to train and “score” the ears from 0% to 100% with that scale.
Guide for scoring how much of the ears were eaten, credit: Anastasia Bodnar
There was no need to have them score images that showed no consumption – or even complete consumption of the corn, so I sorted every experiment into one of three bins as to whether no corn was consumed, some of one or both of the ears were consumed, or both of the ears were fully consumed. Since a lot of time elapsed during the first analysis, we scooped up any more experiments completed on the website after our July deadline. By February they were all sorted, annotated, and ready for Kevin’s scoring team. They hammered through all the images and Kevin scanned their hard-copy data sheets so we could enter them into our spreadsheet.
Next came the careful process of checking the handwriting to make sure no 4s looked like 9s, and that the team followed the directions. I searched for individual bias (e.g. did person #3 tend to rate ears as higher or lower than the average?) and the variation was really low between each person. The fantastic thing about this data was that if the ear actually had 75% of the corn remaining, about half of the team put down 70%, and the other half, 80%, giving us an average that would be very close to the real value. Take one or two team members out and the data hardly changed at all.
Repeatable. But was it reliable? How could I tell that no one in the review team was fudging their numbers to prefer one result over another? They were all blinded. They had no idea which ears were GMO and which were non-GMO.
The blinded leading the blinded
Meanwhile, I sent of random samples of the GMO and non-GMO maize kernels that were collected at the start of the project to be analyzed for purity and composition. We needed to make sure that there wasn’t significant cross-pollination between the two plots that could erode our ability to draw conclusions from this experiment. Moreover, we needed to verify, independently, that all of the genetically engineered traits were present in the GMO variety as promised.
Seed samples labeled with random numbers, ready to ship!
There were supposed to be six different Bt traits and two herbicide-tolerance traits to consider, and if it is true that just one of them made wild animals skittish they all needed to be there. We also needed to make sure that Monsanto sent us comparable varieties of corn. If one had less protein or more starch than the other, that could confound our experiment. The tests came back and showed us that there was no evidence of cross-pollination between the plots, all of the traits were where they needed to be, and the compositions of each variety were the same. When the composition samples were sent off, they were assigned random numbers so the testing lab would not know which was which.
The next step, this spring, was to hand off the data to our statistician, Bill Price, who also joined the project. Bill developed some statistical models to analyze the data that came from Kevin’s team. I assigned new numbers to the genotypes in our data (based on the same random numbers I used for the composition analyses) so that Bill could focus on the data and not think about which one was which. He knew from his experience that even “A” and “B” carried meaning so he wanted random numbers. He got “253297” and “442392”. Can you tell which was which? (At the time of writing even I have to go check the spreadsheet to know.)
Why all the layers of “blinding?” Science is the search for empirical truth – knowledge that can be tested and verified through experimentation. Nature does not care what we believe – it operates the way it will – and our task as scientists is to determine the facts as carefully as we can. Human minds are tricky things, though, and we introduce subtle biases – sometimes without knowing it. Good scientists are committed to determining the truth no matter what their prior beliefs are. Doing experiments blinded ensures that subtle biases cannot creep into the experiment based on how we think the experiment should turn out. We also blinded our participants for the same reason. These details make the results more trustworthy for everyone involved – more reliable – and this is a standard that we hope will be adopted by more scientists on all sides of the biotechnology issue.
Presenting to peers
In June, we had our first opportunity to present our results to the scientific community. I gave a talk at the Plant Biology 2017 conference organized by the American Society of Plant Biologists. The conference was held in Honolulu, and brought scientists from around the country and from across the Pacific to learn about and discuss each others research with talks, poster presentations, and social events. I also organized a science communication workshop at the beginning of the conference. Presenting new research at conferences is an important step on the path to publication because it allows your fellow scientists an opportunity to ask questions, propose analyses you haven’t thought of, and generate a little buzz in the community as well.
Metacorn: Karl wears corn to talk about corn. Credit: Kevin Folta
It can also be a bit of fun. I’ve presented on my thesis research at a maize genetics conference before – the hardest, most nerve-wracking presentation I’ve ever given in my life. But standing in a room of experts, once you realize that you’re the expert on your own research it gets a lot easier. Now add the fact that we’ve got a fun story involving public questions and controversy, Nobel Laureates, funny wild animals, excited kids, and the biggest acknowledgements slide that you’ve probably ever seen. It was a blast! I filmed the talk, so don’t worry – you’ll get to see it. If you were paying attention to our Twitter feeds you may have seen some shots of a slide I prepared for the inevitable news leaks coming out of the conference. I saw cell phone cameras focusing during our big data slides, so I was ready with our social-media-friendly slide right after that. Are you ready?
You heard it here first at #PlantBio17, folks. All completely true. Credit: Jen Robi
Boom. Controversial, I know. The scientists in attendance were in good humor with this, so I proceeded to my next, and most important slide. I wasn’t just there to share our research – I had another agenda: to convince my colleagues that research can also be outreach. Afterward, several people told me that this was the most interesting part. We tend to think of research as discovering knowledge, and outreach as communicating knowledge. I think differently. I think that good research can also be designed as outreach, to both discover and communicate knowledge at the same time. And I believe that we can take this model and apply it to even bigger questions about food, biotechnology, and agriculture. Together, we can change the way that science is done and create a better informed society.
Paper the town
We’re getting down to the last phase of our project. With our results in-hand, and our conference presentation and discussions behind us, we’re busy preparing our paper to submit it to a peer-reviewed journal. For those who are not familiar with this step, we’re carefully writing up the methods, formatting the data, and making decisions on how to best present our results so that everyone can understand them, and so that other scientists could replicate them. There’s more data than what I presented in my talk, including validating our methodology, and so our challenge now is fitting it all together as a cohesive article. There are so many ways to present the data, so we’re choosing what tables, charts, and graphics communicate the most information.
Scientific Squirrels
The ears of corn we used for the experiment were donated by Monsanto, and we signed a Material Transfer Agreement with them, which allowed us to conduct our experiments with their corn, and laid out everyone’s rights. As part of that agreement, we will be sharing our results and our draft of the paper with them prior to submitting it to a peer-reviewed journal.
This is a very standard practice when scientists are studying patented material owned by someone else, and it ensures that we can publish our results – whatever we find – and they get a heads-up on it before we go to publication. Their scientists may even make suggestions such as how to describe their maize varieties or suggest analyses, but we alone have the power to do anything based on those suggestions. To show everyone how this process works, when all is said and done we will show you what we sent them, what they sent back, and any changes we made based on that information.
Then, the paper goes off to a scientific journal to undergo peer review, where anonymous scientists will pick it apart, rate the quality, novelty, newsworthiness, etc. They may recommend publication, revision, or rejection. We might have to make changes and send it back, and we’ll keep you updated on the process when it happens. When we get it published later this year, we’ll have so many stories to tell about this project. It will make a splash!
Credit where credit is due
We can’t take all the credit – because so much of the success of this experiment is due to the work of our Citizen Scientists, donors, and supporters. That’s why we’re taking special care to make sure that everyone involved, from donors to participants, get credit for their contributions.
That’s right, if you were a part of this project, YOU WILL GET YOUR NAME IN OUR PAPER! Although our list is huge, online publishing has no practical size limits so you will get to see your name preserved permanently as part of the scientific record in a supplemental acknowledgements section. To make sure that each and every one of you get credited the way you want, we are sending out a survey over the next couple days to our study participants. Look for it, and please fill it out right away! If you do not receive it by Monday, please contact us and we will make sure that you get it.
Remember when you signed up for the experiment, we agreed to keep your personal information private. So in order to credit you we need your permission – and the great part of this is if you want to highlight the contributions of your K-12 research team, you can choose how to credit them. If you want to remain anonymous, you can choose that option as well. (Experiment.com donors who did not participate in the experiment do not need to fill out this survey as their contribution is already public and will be included in our acknowledgements.) As an extra incentive for finishing our survey, we’re going to award some prizes for participation.
Did you say awards? Prizes?
Yes I did! Remember how I said that I read through over 2,000 observations? How I sifted through hundreds of experiments, and all the images they contained? I could see all the work you put into these experiments. I could sense the enthusiasm, the creativity, the frustration, and the satisfaction. I remember one citizen science team that kept repeating their experiment over and over again, taking notes saying that no animals touched the corn. Diligently, for two weeks, taking photos, resetting the experiment, and gathering weather data. Then came the day that they finally attracted the attention of wild animals, corn was getting eaten and exclamation points were everywhere. I half jumped out of my seat! That kind of dedication deserves recognition.
More teams impressed me with their detail-oriented approach – taking careful and comprehensive observations. Others took incredible and sometimes hilarious photos. There was some good humor involved in some of the experimental data – “kids” in the list of wild animals active in the area, signs posted in the background of their experiments, and prosaic observations. I could see some future scientists in there. We need an award ceremony.
When you get the survey, fill it out and you are entered to win one of several awards for your participation, including several randomly-selected prizes. Fill it out by August 25th!
When we submit our paper we will hold a live streamed broadcast to announce the submission, release some of our results to the public, and celebrate everyone’s contributions and announce the awards. Stay tuned, because the most exciting part is yet to come.
I’m Alma Laney from The Mad Virologist, and I work with plant viruses and the insects that help them spread. I’m happy to announce that Biology Fortified just published a new infographic that I helped develop with Layla Katiraee and the rest of the team here at Biology Fortified to help explain how virus resistance is created in genetically engineered crops.
The GMOs Revealed infographic on Virus Resistance distills a lot of information about how to engineer this trait into plants, with papayas and squash as the current examples on the market. There is really a lot more to this issue than these two examples, so I wanted to give a detailed overview explaining the challenges that farmers face with viral plant diseases, methods of control, and approaches to engineering resistance. Go to the infographic page for the short version, but if that’s not enough to ‘inoculate’ your curious mind, read on!
Introduction
Plant viruses can be serious pathogens in crops as they can cause anywhere from minor losses to a total loss. Viruses can infect crops in a number of ways ranging from being transmitted by contaminated tools, seed and pollen infections, infection of tubers or other vegetatively propagated material, and by arthropod vectors1 (what mites and insects are collectively known as). Most plant viruses are transmitted to crops via arthropod vectors. Plant viruses are of concern to farmers because once they get into a crop, all you can do is try and prevent their spread. Because of this, control strategies focus on preventing the introduction and spread of these viruses.
Control of plant viruses
There are several strategies that farmers use to prevent and control viral infections:
Papaya leaf infected with PRSV, Thailand. Credit KJHvM
Use certified seed or plants, which have been tested for known pathogens. Lots containing pathogens are rejected by the company.
Control weeds around the fields as weeds can harbor both viruses and their vectors, serving as a source of inoculum for the field.
Limit the spread of soil and the use of dirty implements. Some plant viruses can be carried by infested soil and contaminated implements can transfer the virus to healthy plants. There are many disinfectants that can be used including dilute bleach and milk solutions.
Use seed treatments and/or spraying insecticides on the crop. Since most plant viruses are transmitted by arthropod vectors, this can be an effective strategy but it does not work in cases where the viruses are transmitted quickly (in non-persistent or semi-persistent transmission; see Table 1).
A control strategy that isn’t always effective is to use a mild strain of a virus that is inoculated onto a plant. The mild strain can induce resistance to more severe strains, but this is problematic as mixed infections with other viruses can cause severe disease, the mild strain could become more severe, the mild strain would still cause economic losses, or the mild strain just may not work. This strategy is called cross protection.
Once a virus is in a field, farmers can take action by roguing symptomatic plants (removal of infected plants followed by destroying them). This can be a costly measure and is used in cases where the type of virus can be transmitted quickly.
Table 1: The types of virus transmission2 detailing how long it takes to acquire a virus, how long it takes before the virus can be transmitted, and an example of each.
Transmission type
Virus acquisition
Virus transmission
Example
Non-persistent
As little as a few seconds
A few minutes
Papaya ringspot potyvirus
Semi-persistent
Several minutes
Minutes to hours
Cauliflower mosaic caulimovirus
Circulative, non-propagative
Minutes to hours
Hours to days
Barley yellow dwarf luteovirus-PAV
Circulative, propagative
Minutes to hours
Up to a few weeks
Tomato spotted wilt tospovirus
However, one of the best strategies is to engineer resistance to plant viruses. There are several examples of plant virus resistance genes that can be found in the germplasm (a collection of seed and/or plant tissue reflecting a variety of genotypes). However, for some crops, there are no known resistance genes or if they exist, they are found in wild relatives and attempts to introgress (move a trait into a crop plant by conventional breeding) the genes have not been successful. This is where genetic engineering can provide a solution. Genetically engineered plant virus resistance induces two different forms of resistance and one of the methods can use two different approaches.
Transgenic approach to virus control
Early on, virologists transformed plants with the complete virus coat protein gene, which forms the shell of the virus to protect the genetic material. It was found in the case of Tobacco mosaic tobamovirus (TMV), that over-expression of the coat protein gene led to virus resistance because the excess coat protein interfered with the ability of the virus to complete its lifecycle and move systemically in the plant3. Based on this, transgenics for other viruses were generated including Papaya ringspot potyvirus (PRSV), Cucumber mosaic cucumovirus (CMV), Zucchini yellow mosaic potyvirus (ZYMV), and Watermelon mosaic potyvirus (WMV). However, later research uncovered a second way that transgenics can induce resistance to plant viruses: triggering the RNA silencing (RNAi) pathway found in plants.
The discovery of RNAi was revolutionary. It has led to other control strategies as well as providing a powerful tool for functional analysis of genes. In plants, the RNAi pathway serves as a type of immune system for plants to target pathogens4, including viruses. There are multiple ways that the RNAi pathway can be triggered:
Predicted folding structure of the coat protein gene for a Hawaiian isolate of Papaya ringspot potyvirus (GenBank Accession # S46722.1) generated by A.G. Laney using mFold. Note, this is only one of many predicted folding structures for this gene and is used to illustrate that there are double stranded regions of the gene that could trigger the RNAi pathway.
dsRNA: Most of the described plant viruses have RNA genomes and one of the byproducts of viral replication is double-stranded RNA (dsRNA)5. This dsRNA is very stable and triggers the RNAi pathway. However, even with DNA viruses, some of the genes overlap and can trigger the RNAi pathway too6. With the transgenics that utilized the complete coat protein but triggered the RNAi pathway, it was found that in some cases the secondary structure of the RNA produced regions with dsRNA that could trigger RNA silencing in plants (see image at right)7.
Short hairpins: The discovery that short dsRNA segments could trigger RNAi led to the use of constructs that generate a hairpin cassette that forms into dsRNA8. Early uses of virus-derived transgenic resistance used entire genes; however, with advances in our understanding of the RNAi pathway, many researchers have adopted the hairpin cassette method as it allows for the targeting of multiple viruses and/or viral genes9. This has several benefits. By targeting multiple genes, researchers are able to minimize the chances of the targeted virus developing resistance to the transgenic plant. The hairpin cassette technique also allows researchers to target multiple viruses with a single construct.
Virus-derived transgenic products on the market
Virus Resistant Papaya, credit: KJHvM
The most widely known transgenic on the market is papaya that is resistant to PRSV10. This product actually saved the papaya industry in Hawaii as PRSV makes fruit unmarketable and eventually kills infected trees. Complicating matters, PRSV is transmitted by several aphids non-persistently (it’s carried in the stylet of the aphid) so it transmits as soon as the aphid probes the plant tissue. Because of this, attempts to control the aphid vectors by spraying does not work as the virus has already been transmitted by the time the aphids are killed.
Resistance to the virus was the only option; however, although resistance to PRSV has been identified in wild relatives, all attempts to introgress the trait were have not been successful introduced into cultivated papaya from a wild relative until just recently11 after 50 years of attempts by plant breeders. The only strategy that papaya growers could use was moving their operations to another island. Each time they moved, there was a short reprieve, but the virus eventually made it to that area. To combat this, work was started to investigate the potential for cross protection; however, it was not effective with the isolates found in Hawaii. Luckily at around that time, news of a new technique was announced that used a transgenic with the coat protein of a virus that provided resistance to that virus. So work on a transgenic began. By the time PRSV made it to the last papaya growing area in Puna, Hawaii, the transgenic was ready and the industry was saved.
There are two different transgenic events for virus resistance in summer squash. The first, ZW-2012, targets ZYMV and WMV whereas the second, CZW-313, targets CMV in addition to ZYMV and WMV. ZYMV and WMV are related to PRSV (they are all in the same genus, Potyvirus) and are targeted by introduction of the entire coat protein gene that induces the RNAi pathway in plants. The way that the CMV construct works is interesting. It seems that the introduction of the coat protein gene acts by interfering with the life cycle of the virus and by inducing the RNAi pathway.
Virus-derived transgenic products in development
There are a number of transgenic crops that utilize virus-derived resistance currently in development. These range from rice plants engineered to resist Rice grassy stunt tenuivirus14 to cotton plants that are resistant to Cotton leaf curl Kokhran begomovirus15 to tomatoes resistant to Tomato yellow leaf curl begomovirus16 to lettuce that is resistant to Mirafiori lettuce big-vein ophiovirus17. (See here for an article on Biofortified about virus-resistant black beans in Brazil.) Each of these viruses causes severe losses in their respective crops and durable resistance either has not been found or has been hard to introduce into the respective crop plants. This is just a small subset of the virus-derived transgenic plants that are in various stages of development worldwide. However, there are several viral diseases that cause food insecurity and/or severe economic losses that have transgenic solutions. Two that are threats to food security will be discussed below:
Cassava mosaic disease (CMD)
VIRCA Plus director Dr. Nigel Taylor working with Cassava. Credit: Donald Danforth Plant Science Center
This disease contributes to food insecurity in Africa and Southern Asia18 and is caused by at least 10 related virus species in the genus Begomovirus. Losses to CMD are due to the absence of tubers on infected plants. Since cassava grows well in poor soil with less rain than other staple crops, it is widely grown in sub-Saharan Africa. Because of the severity of the losses and because cassava is often a crop counted on to help reduce the effects of famine, it is an ideal target for virus-derived transgenic resistance.
By targeting a gene that is essential for begomovirus replication (AC1), researchers were able to generate cassava plants that were resistant19 to African cassava mosaic begomovirus as well as two related cassava begomoviruses. This was then improved further by targeting two additional genes essential for begomovirus replication (AC2 and AC3)20. The effort to develop virus-derived transgenic resistance to CMD has been included in the VIRCA Plus project which combines resistance to CMD, cassava brown streak disease and nutritional improvements (transgenic fortification with zinc and iron) with to cassava.
Cassava brown streak disease (CBSD)
Like CMD, CBSD is a threat to food security. Although CBSD can reduce tuber size it does not result in no tubers as with CMD. However, what remains of CBSD symptomatic tubers are inedible due to necrosis in the tubers making infected crops a near total loss21. CBSD is caused by viruses that are transmitted by whiteflies. However, there are only two known species, Cassava brown streak ipomovirus and Ugandan cassava brown streak ipomovirus, and both are generally just referred to as CBSV. Like CMD, the viruses that cause CSBD have been targeted using virus-derived transgenic resistance22 and as mentioned above, transgenic resistance to these viruses have been included in the VIRCA Plus project.
Other avenues for using RNA silencing
The usefulness of RNA silencing is not limited to resistance to plant viruses. There have been other examples of this technology being used to prevent browning due to oxidation (Arctic apple and the Simplot Innate potatoes – see articles on Biofortified about the apple and potato) and reducing acrylamide formation in cooked potatoes (also the Innate potato). Another area being explored that relates to transgenic virus resistance is the development of RNA silencing for arthropod vectors of plant viruses. This is still in the early stages of development, but shows great promise in offering more options for farmers to use in integrated pest management. As with virus-derived resistance, RNA silencing for insect control has focused on turning off genes that are essential for the insect to live, such as v-ATPase subunit A in whiteflies23, or genes that are essential for insect-plant interactions, such as C002 which is expressed in the salivary glands of aphids24. There will be an additional post discussing the use of RNA silencing for other uses in the near future.
Conclusions
Virus-derived transgenic resistance holds great promise in sparing growers and consumers the costs of losses due to virus infection. Furthermore, this technology has saved at least one crop, papaya grown in Hawaii, and holds the potential to grant those in developing nations food security by preventing losses in staple crops. Some of the other benefits of this approach to controlling plant viruses is that it reduces sprays that were used to control the arthropod vectors, while not altering how the crops are grown. One of the main challenges is that resistance to one strain of virus may not give strong resistance to other strains, so the evolution of new virus strains must be closely monitored.
References:
Leitner et al., 2015. Arthropod Vectors and Disease Transmission: Translational Aspects. PLoS Neglected Tropical Pathogens 9(11): e0004107. DOI: 10.1371/journal.pntd.0004107
Whitfield et al., 2015. Insect vector-mediated transmission of plant viruses. Virology Volumes 479–480: 278–289 DOI: 10.1016/j.virol.2015.03.026
Beachy, 1999. Coat-protein-mediated resistance to tobacco mosaic virus: discovery mechanisms and exploitation. Philos Trans R Soc Lond B Biol Sci 354:659-664. DOI: 10.1098/rstb.1999.0418
Obbard et al., 2009. The evolution of RNAi as a defence against viruses and transposable elements. Philos Trans R Soc Lond B Biol Sci 364(1513): 99–115. DOI: 10.1098/rstb.2008.0168
Weber et al., 2006. Double-Stranded RNA Is Produced by Positive-Strand RNA Viruses and DNA Viruses but Not in Detectable Amounts by Negative-Strand RNA Viruses. Journal of Virology. 80(10): 5059–5064. DOI: 10.1128/JVI.80.10.5059-5064.2006
Li et al., 2014. Suppression of RNA Silencing by a Plant DNA Virus Satellite Requires a Host Calmodulin-Like Protein to Repress RDR6 Expression. PLoS One. DOI: 10.1371/journal.ppat.1003921
Lindbo and Falk, 2017. The Impact of “Coat Protein-Mediated Virus Resistance” in Applied Plant Pathology and Basic Research. Phytopathology 107(6): 624-634 DOI: 10.1094/PHYTO-12-16-0442-RVW
Jia et al., 2007. A strategy for constructing and verifying short hairpin RNA expression vectors. J RNAi Gene Silencing 3(1): 248–253. PMCID: PMC2737214
Lambeth et al., 2010. A direct comparison of strategies for combinatorial RNA interference. BMC Molecular Biology 11:77. DOI: 10.1186/1471-2199-11-77
Gonsalves et al., 2004. Transgenic Virus Resistant Papaya: From Hope to Reality for Controlling Papaya Ringspot Virus in Hawaii. APSnet Features. Online. DOI: 10.1094/APSnetFeature-2004-0704
Siar et al., 2011. Papaya ringspot virus resistance in Carica papaya via introgression from Vasconcellea quercifolia. Euphytica 181: 159-168 DOI: 10.1007/s10681-011-0388-z
Fuchs and Gonsalves, 1995. Resistance of Transgenic Hybrid Squash ZW-20 Expressing the Coat Protein Genes of Zucchini Yellow Mosaic Virus and Watermelon Mosaic Virus 2 to Mixed Infections by Both Potyviruses. Nature Biotechnology 13: 1466 – 1473 DOI: 10.1038/nbt1295-1466
Tricoll et al., 1995. Field Evaluation of Transgenic Squash Containing Single or Multiple Virus Coat Protein Gene Constructs for Resistance to Cucumber Mosaic Virus, Watermelon Mosaic Virus 2, and Zucchini Yellow Mosaic Virus. Nature Biotechnology 13: 1458 – 1465 DOI: 10.1038/nbt1295-1458
Shimizu et al., 2013. Strong Resistance Against Rice grassy stunt virus Is Induced in Transgenic Rice Plants Expressing Double-Stranded RNA of the Viral Genes for Nucleocapsid or Movement Proteins as Targets for RNA Interference. Phytopathology 103: 513-519 DOI: 10.1094/PHYTO-07-12-0165-R
Yasmeen et al., 2016. Amplicon-Based RNA Interference Targeting V2 Gene of Cotton Leaf Curl Kokhran Virus-Burewala Strain Can Provide Resistance in Transgenic Cotton Plants. Molecular Biotechnology 58: 807-820 DOI: 10.1007/s12033-016-9980-8
Fuentes et al., 2016. Field Trial and Molecular Characterization of RNAi-Transgenic Tomato Plants That Exhibit Resistance to Tomato Yellow Leaf Curl Geminivirus. Molecular Plant-Microbe Interactions 29: 197-209 DOI: 10.1094/MPMI-08-15-0181-R
Kawazu et al., 2016. Development of marker-free transgenic lettuce resistant to Mirafiori lettuce big-vein virus. Transgenic Research 25: 711-719 DOI: 10.1007/s11248-016-9956-2
Alabi et al., 2011. Cassava Mosaic Disease: A Curse to Food Security in Sub-Saharan Africa. APSnet Features. Online. DOI: 10.1094/APSnetFeature-2011-0701
Chellappan et al., 2004. Broad Spectrum Resistance to ssDNA Viruses Associated with Transgene-Induced Gene Silencing in Cassava. Plant Molecular Biology 56: 601-611 DOI: 10.1007/s11103-004-0147-9
Zhang et al., 2005. Resistance to cassava mosaic disease in transgenic cassava expressing antisense RNAs targeting virus replication genes. Plant Biotechnology Journal 3: 385-397 DOI: 10.1111/j.1467-7652.2005.00132.x
Patil et al., 2015. Cassava brown streak disease: a threat to food security in Africa. Journal of General Virology 96: 956-968 DOI: 10.1099/vir.0.000014
Ogwok et al., 2012. Transgenic RNA interference (RNAi)-derived field resistance to cassava brown streak disease. Molecular Plant Pathology 13: 1019-1031 DOI: 10.1111/j.1364-3703.2012.00812.x
Thakur et al., 2014. Enhanced Whitefly Resistance in Transgenic Tobacco Plants Expressing Double Stranded RNA of v-ATPase A Gene. PLoS One 9(3): e87235 DOI: 10.1371/journal.pone.0087235
Pitino et al., 2011. Silencing of Aphid Genes by dsRNA Feeding from Plants. PLoS One 6(10): e25709 DOI: 10.1371/journal.pone.0025709
Written by Guest Expert
Alma Laney works with plant viruses and the arthropods who vector them and on his blog The Mad Virologist he covers all aspects of virology from human pathogens to archaea viruses and everything in between. http://themadvirologist.blogspot.com/
Source: Portal Educativo. Image slightly modified.
Biotechnology is poised to become one of the most valuable scientific revolutions of the 21st century. Because the field is developing so quickly, the gap between expert and non-expert knowledge is increasing at a time when societal decisions about it are becoming more and more important. So how do we promote biotechnology literacy in the classroom? What should non-experts know about genetic technologies in order to make informed decisions? I conducted a study to answer these questions, and here is what I found.
Even though scientific knowledge is an important part of science literacy, how people feel about a technology – their general positive or negative attitudes – also plays a role in their decision-making. In fact, there’s evidence that attitudes play a greater role than knowledge in determining students’ behavior toward biotechnology. I set out to understand what issues undergraduate students draw upon when they reason about genetic technologies. I also wanted to know whether classroom dialogue about biotechnology influences their attitudes and understanding. This information can provide a window into the conceptual frameworks that students use to make decisions about genetic technologies, and can help educators and communicators develop specific strategies for connecting with their audiences.
Students discuss biotechnology
Twenty years ago, my postdoctoral mentor Pamela Ronald launched an innovative course designed for non-science majors at UC Davis. Genetics and Society engages students in the science, politics, social issues, ethics, and economics surrounding biotechnology. It remains popular today. Recognizing the importance of dialogue around this complex topic, Pam introduced “discussion sections” into the course. During the discussion sections, students engage in rational discourse about a biotechnology issue – for example, whether or not all food containing genetically engineered (GE) ingredients should be labeled as “GMO”. The discussion sections provide an opportunity for students to share their thoughts and consider the many facets involved in decision-making about biotechnology. Scientific arguments used in the class are required to be evidence-based, and students are graded on the credibility of their sources. While students in Genetics and Society generally enjoy these peer-to-peer discussions, no one had looked closely at how they influence their understanding and attitudes about genetic technologies.
At the beginning of the course, I asked the students to state their attitudes on seven different biotechnology applications. Three topics related to food: whether or not we should label GMOs, whether GE of plants should be prohibited, and whether GE of animals should be prohibited. I also asked the students to justify their attitude for each topic. At the end of each weekly discussion section, during which a group of students presented on an individual application/topic, I collected this information a second time from the students in the audience. These pre-post attitudes with corresponding reasoning provided the data for my study. Figure 1. Student attitudes significantly changed for three topics following classroom dialogue. Source: Anderton & Ronald, Journal of Biological Education, 2017.
I started by looking for significant changes in students’ attitudes following the discussion sections. I found significant changes for three topics: GMO labeling, GE of animals, and the FDA ban on 23andme’s health reports* (Figure 1). Because the students did not appear to have familiarity with the 23andme ban at the beginning of the course, I didn’t select that topic for further analysis. In the end, I selected GMO labeling and GE of animals, as well as two topics for which I didn’t observe significant attitude changes (DNA fingerprinting and human embryo editing research) for further analysis. Pam and I reasoned that it was important to take a close look at student reasoning in the presence and absence of attitude changes, because learning can happen even if a person doesn’t change their mind.
Analyzing the themes
Using the justifications given by the students for their attitudes on the four topics selected above – GMO labeling, GE of animals, DNA fingerprinting and human embryo editing – I performed an approach called thematic analysis, in which I looked for overarching patterns or themes that were prevalent in students’ reasoning about biotechnology. Through an iterative process, I identified seven major themes that students drew upon in their justifications (Figure 2). I also tallied the number of times I detected a change in the use of a theme following a given discussion section (i.e., whenever a student adopted new reasoning or abandoned prior reasoning following a discussion section). Figure 2. Seven overarching themes related to student reasoning about biotechnology. Source: Anderton & Ronald, Journal of Biological Education, 2017.
Our preliminary evidence suggests that the discussion sections – and perhaps classroom discourse in particular – provide students with a more nuanced understanding of biotechnology. For example, students generally increased their use of “Middle Way” reasoning following the discussion sections. This suggests that they developed a greater appreciation of regulations that consider biotechnology applications on a case-by-case basis. We also observed increased use of the Economic theme following the two discussion sections for which students had significant attitude changes. It is possible that the economic considerations of genetic technologies can sway people’s attitudes, but this remains to be proven.
Scientific decision-making involves more than just facts. By better understanding the complex processes that take place when students learn and make decisions about genetic technologies – like we did in this study – educators can connect with their audiences and promote biotechnology literacy. A more informed and nuanced discussion will help our society determine the best ways to use biotechnology and to direct our focus as it continues to evolve.
To access the full version of the manuscript, Hybrid thematic analysis reveals themes for assessing student understanding of biotechnology, please go to: http://dx.doi.org/10.1080/00219266.2017.1338599.
Please address any questions or comments to Brittany Anderton at bnanderton@udavis.edu.
*The FDA banned 23andme genetic health reports in November 2013, citing concerns about the accuracy and usefulness of such information to consumers. In October 2015 the ban was lifted, and 23andme resumed offering carrier-status testing, though no longer offers testing for health conditions such as cancer and heart disease.
Written by Guest Expert
Brittany Anderton seeks to improve the intersection of science and society by educating the next generation of responsible scientists and citizens. She has a PhD in cancer biology, studied the teaching and communication of biotechnology as a postdoctoral fellow at UC Davis, and is now the Associate Director of Research Talks at iBiology and Lecturer at CSU Sacramento.